What's in a name? Taxonomists arguing over the finer points of speciation could give you an earful on this. On a somewhat related note, I once heard a person exclaim, "When I see a monkey give birth to a man, I'll believe in evolution." They were probably quoting or paraphrasing some famous ignoramus, and the who of the matter is merely trivial. The point is that, for want of education, there are many who simply do not understand that modern humans did not descend from modern apes, but rather that both groups are descended from a common ancestor.
I can attest from personal experience that, for those with no education in biology past the age of eighteen (formal or informal - I mean those who do not even casually look into the subject), it can be extremely difficult to communicate this point in an intelligible and accurate manner. We are, after all, talking about long-extinct transitional forms creating a chain going back in time and forward again down multitudinous paths to cement the cousinship of all life on the planet. To the uninitiated and skeptical, this can be a tough concept to wrap one's mind around! The metaphor of a growing tree, with the very tips of each twig representing extant life, can be a useful one. But there is a real life example staring us in the face which can help drive the concept home, and that example is ring species.
Here is a general example of a ring species: imagine three groups of animals, A, B, and C. Animals of group A interbreed with animals of group B, and animals of group B interbreed with animals of group C - but none of the animals of group A interbreed with any of the animals of group C. The interesting part is where we see this in living animals.
An example of this phenomenon is in the Larus gulls of the Arctic Circle. In Europe live two species of gull, the Herring gull and the Lesser Black-backed gull. These two groups do not interbreed, though they often perform their mating in mixed groups (that is, all together at the same time, but Herring gulls mating with Herring gulls only and Lesser Black-backed gulls mating with Lesser Black-backed gulls only). However, the Herring gulls of the UK will interbreed with the Herring gulls of North America, which look kind of like them. The American Herring gull will also interbreed with the Vega Herring gull of Northern Russia, which kind of looks like it. Vega's gull interbreeds with Birula's gull, and the two also look somewhat similar to each other. Birula's gull interbreeds with Heuglin's gull, which in turn interbreeds with the Siberian Lesser Black-backed gull, which in turn interbreeds with - did you guess it yet? - the Lesser Black-backed gull of Europe! Here it is in graphic format:
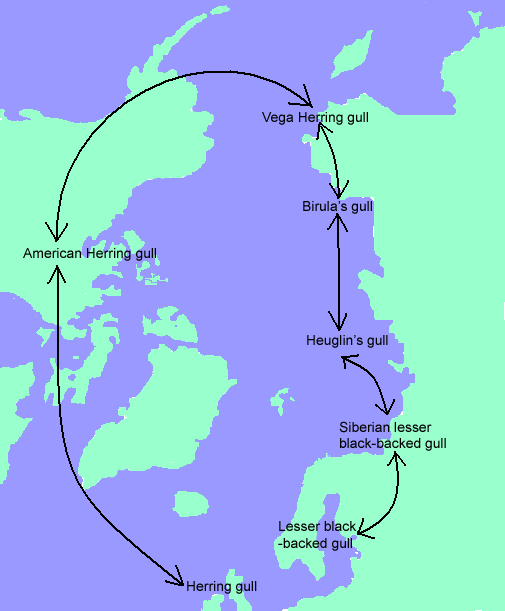
{kind=link}
So, to recap: the Herring gull and Lesser Black-backed gull do not interbreed with each other, yet each interbreeds with geographic neighbors in a continuous ring which connects the non-interbreeding groups.
A more striking example is in the Ensatina salamanders of Western North America. In the mountains surrounding the Central Valley of California, the Ensatina accomplishes in one state what the Larus gulls do all around the Arctic Circle. Some nineteen species of salamander live in areas forming a vague loop around the Central Valley, and at the Southern end, near Mexico, this loop is "clipped" (as the Arctic loop above is "clipped" in Europe). To the West lives the Monterey Ensatina, Ensatina eschscholtzii eschscholtzii; to the East, the Large-blotched Salamander, Ensatina eschscholtzii klauberi. Hearkening back to our earlier, general example, the Monterey Ensatina is our "A" species, and the Large-blotched Salamander our "C." All the other seventeen species of Ensatina living around the loop are "B" species: capable of interbreeding with each other, as well as with A & C.
Interbreeding (more precisely, the ability by interbreeding to produce viable offspring) is often held to be the gold standard of taxonomic distinction: if you interbreed, you're probably the same species; if you don't, you're definitely not the same species. One would think that such a binary, cut-and-dry system would be foolproof, but Mother Nature is as cunning as she is complicated. Ring species, by their very existence, present a nigh-insoluble paradox to this view - if A & B are the same species, and B & C are the same species, then A & C should be the same species; yet A & C do not interbreed, and so by our definition of "species," it would appear that we cannot resolve our inconsistent desires to at once classify B with each of A and C, but also separate A and C from each other taxonomically.
There is, of course, a third option: the idea of distinct "species" is fundamentally illusory, and our taxonomy (while useful) is merely a language of convenience. This, as it turns out, is the case! In The Ancestor's Tale, Richard Dawkins explains these examples and points out just exactly how instrumental they can be to understanding how speciation naturally occurs:
Ring species like the salamanders and the gulls are only showing us in the spatial dimension something that must always happen in the time dimension. Suppose we humans, and the chimpanzees, were a ring species. It could have happened: a ring perhaps moving up one side of the Rift Valley, and down the other side, with two completely separate species coexisting at the southern end of the ring, but an unbroken continuum of interbreeding all the way up and back round the other side. If this were true, what would it do to our attitudes to other species? To apparent discontinuities generally? (pg. 303)In the terms of our general example, some B species could be considered a distant common ancestor to extant species A and C - when no B species is present, A and C appear to be distinct (and for good reason). But when we see B alongside A and C, the line separating the two becomes fuzzier as their kinship is thrust into our faces. Alternatively, A could be the ancestral species, B the transitional forms, and C the extant species. At no point does A "turn into" C; rather, it goes through various B stages before arriving, smoothly, at C (which in turn is merely a transitional form "on the way to" D, E, F, and so on). The time scales involved in such transformations, though, are typically on the order of hundreds of thousands of years, and as such are difficult to point to conclusively. Well, it's easier with bacteria, but that opens a whole 'nother can of bees.
Later, Doctor Dawkins lays bare the philosophical underpinnings of such problems as they relate to modern taxonomy (and here I thought the philosophy was my job!):
Ernst Mayr, distinguished elder statesman of twentieth-century evolution, has blamed the delusion of discontinuity - under its philosophical name of Essentialism - as the main reason why evolutionary understanding came so late in human history. Plato, whose philosophy can be seen as the inspiration for Essentialism, believed that actual things are imperfect versions of an ideal archetype of their kind. Hanging somewhere in ideal space is an essential, perfect rabbit, which bears the same relation to a real rabbit as a mathematician's perfect circle bears to a circle drawn in the dust. To this day many people are deeply imbued with the idea that sheep are sheep and goats are goats, and no species can ever give rise to another because to do so they'd have to change their 'essence'.Dawkins refers to this "essentialist" viewpoint as part of "the tyranny of the discontinuous mind." Discontinuity is necessary in order to make classification coherent in the first place (which is good, as classification is often useful), but it is often the case that such discontinuities aren't actually present - at least, not at the levels our minds are evolutionarily prepared to deal with. Under the thumb of a discontinuous mind, it is easy to see why the fiction of a six-thousand-years-past Creation of the world and all its "distinct" species is so damned attractive to so many: it fits into our tiny, tiny brains, with all their genetically programmed instincts. The reality of the situation, though - that speciation is due only to extinction, and all the world is kin - is far subtler, far grander, and far more interesting than any fiction ever dreamt by humankind.
There is no such thing as essence.
No evolutionist thinks that modern species change into other modern species. Cats don't turn into dogs or vice versa. Rather, cats and dogs have evolved from a common ancestor, who lived tens of millions of years ago. If only all the intermediates were still alive, attempting to separate cats from dogs would be a doomed enterprise, as it is with the salamanders and the gulls. Far from being a question of ideal essences, separating cats from dogs turns out to be possible only because of the lucky (from the point of view of the essentialist) fact that the intermediates happen to be dead. Plato might find it ironic to learn that it is actually an imperfection - the sporadic ill-fortune of death - that makes the separation of any one species from another possible. (pg. 308)
No comments:
Post a Comment